
Au-delà des gènes
Les mécanismes de l'épigénétique




Pour qu'un gène soit transcrit, il faut que des facteurs de transcription ainsi que l'ARN polymérase puissent se lier à l'ADN. Seulement, des facteurs épigénétiques régulent l'expression des gènes en les rendant accessibles ou bien inaccessibles à l'ARN polymérase. On parle d'expression ou de répression des gènes.
Deux mécanismes entrent en jeu :
-
La méthylation de l'ADN
-
La modification des histones
La méthylation de l'ADN :
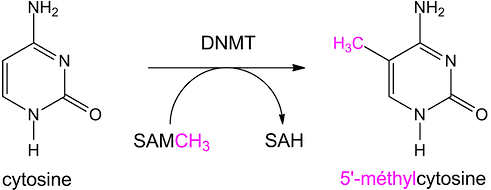
La méthylation se résume donc à l'ajout de groupements méthyle (CH3) par l'intermédiaire d'enzymes particulières telles que l'ADN méthyltransférase (DNMT). Ces enzymes catalysent le transfert d'un groupement méthyle d'une S-adénosylméthionine (SAM) sur des résidus cytosine. La S-adénosylméthionine est un métabolite présent dans les cellules qui agit comme coenzyme dans les transferts de groupements méthyle.
Il existe trois familles d'ADN méthyltransférases ( DNMT ) :
-
DMNT1 : qui maintient la méthylation sur les deux brins d'ADN lors de la réplication.
-
DNMT2 : dont le rôle reste encore incertain.
-
DNMT3 : permettant l'ajout de nouvelles marques de méthylation sur l'ADN.
La méthylation de l'ADN semble être un mécanisme réprimant l'expression des gènes. C'est un processus réversible dont les termes de déméthylation ne sont pas encore prouvés.
Les histones :
Les histones sont des protéines autour desquelles l'ADN s'enroule. Ils ont deux domaines :
-
un domaine globulaire qui forme les contacts avec les autres histones et avec l'ADN.
-
un domaine formant une queue, chargé positivement et qui interagit avec les charges négatives des groupements phosphates du squelette de l'ADN. Ce domaine peut subir des modifications épigénétiques qui vont affecter l'expression des gènes en les réprimant ou en les rendant encore plus accessibles. Toutes ces modifications définissent le code histone.
Les modifications au niveau des histones vont engendrer une changement de la conformation de l'ADN. En effet si des marques épigénétiques jouent un rôle dans la formation d'hétérochromatine, l'ADN sera moins accessible pour l'ARN polymérase et la transcription sera donc impossible. Selon le niveau d'hétérochromatine la transcription aura plus ou moins lieu ce qui régule toute l'expression de nos gènes et donc tout nos caractères. Les modifications des histones est alors indispensable pour la régulation de notre organisme.
On va parler de 4 des modifications des histones :
-
La méthylation :
Elle peut subvenir sur deux acides aminés particuliers, la lysine, méthylée par les protéines d'un certain domaine SET. Cette méthylation joue un rôle de formation de l'hétérochromatine, l'ADN est donc condensé et illisible. La méthylation de la lysine réprime alors l'expression des gènes.
Ainsi que l'arginine méthylée par les PRMT (protein arginine methyltransferase) ayant elle un effet positif sur la transcription.
-
L'acétylation :
C'est l'ajout d'un groupement CH3CCO. L'acétylation se fait sur l'acide aminé lysine par l'intermédiaire de co-activateurs transcriptionnels, les histones acétyltranférases (HAT). On les classe selon deux familles, les MYST et les GNAT. L'ajout de ce groupement acétyle va neutraliser la charge positive de l'acide aminé et modifier la taille de la chaine latérale du résidu de lysine. Cela va déstabiliser la structure de la chromatine en la rendant plus flexible et donc permettre une meilleure transcription des gènes.
Il existe également des enzymes désacétylases qui vont venir détacher les groupements acétyle des histones rétablissant la structure de la chromatine et donc réprimant la transription.
-
La phosphorylation :
C'est l'ajout d'un groupement phosphate PO3 2-. La phosphorylation est catalysée par des protéines kinases qui sont des enzymes catalysant le transfert d'un phosphate vers la chaîne latérale des acides aminés ayant une fonction alcool. Les enzymes phosphatases sont des facteurs qui se chargent de la déphosphorylation.
-
L'ubiquitination :
C'est l'ajout d'une ou plusieurs protéines d'ubiquitine sur des lysines. Trois classes d'enzymes permettent la mise en place de l'ubiquitination, l'E1-ligase, l'E2-ligase et l'E3-ligase. Tandis que les enzymes appelées Ubiquitin Specific Peptidase (USP) catalysent la réaction inverse et permettent la désubiquitination des protéines.
La queue étant chargée positivement et l'ADN négativement, alors ils sont attirés l'un vers l'autre.
L'acétylation a neutralisé la charge positive des queues des histones, l'ADN est plus libre et donc mieux transcrit.
La méthylation de l'ADN ne s'effectue que sur des résidus Cytosine précédant un résidu Guanine. On parle de Dinucléotide CpG.
La méthylation aboutie selon la réaction :